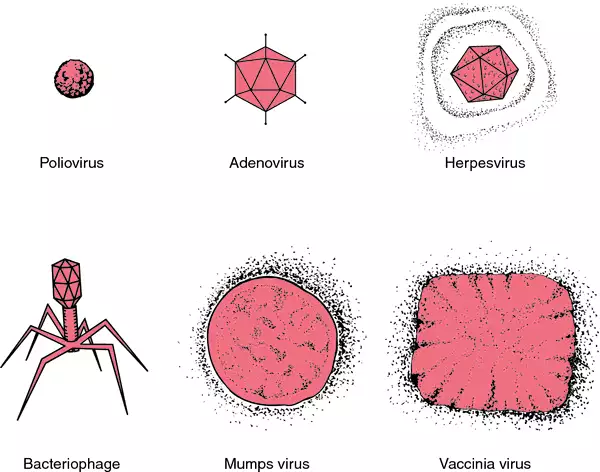
A DNA virus is a virus that has DNA as its genetic material and replicates using a DNA-dependent DNA polymerase. The nucleic acid is usually double-stranded DNA (dsDNA) but may also be single-stranded DNA (ssDNA). DNA viruses belong to either Group I or Group II of the Baltimore classification system for viruses. Single-stranded DNA is usually expanded to double-stranded in infected cells. Although Group VII viruses such as hepatitis B contain a DNA genome, they are not considered DNA viruses according to the Baltimore classification, but rather reverse transcribing viruses because they replicate through an RNA intermediate. Notable diseases like smallpox, herpes, and chickenpox are caused by such DNA viruses.
Group I: dsDNA viruses

Genome organization within this group varies considerably. Some have circular genomes (Baculoviridae, Papovaviridae and Polydnaviridae) while others have linear genomes (Adenoviridae, Herpesviridae and some phages). Some families have circularly permuted linear genomes (phage T4 and some Iridoviridae). Others have linear genomes with covalently closed ends (Poxviridae and Phycodnaviridae).
A virus infecting archaea was first described in 1974. Several others have been described since: most have head-tail morphologies and linear double-stranded DNA genomes. Other morphologies have also been described: spindle shaped, rod shaped, filamentous, icosahedral and spherical. Additional morphological types may exist.
Orders within this group are defined on the basis of morphology rather than DNA sequence similarity. It is thought that morphology is more conserved in this group than sequence similarity or gene order which is extremely variable. Three orders and 31 families are currently recognised. A fourth order â€" Megavirales â€" for the nucleocytoplasmic large DNA viruses has been proposed. Four genera are recognised that have not yet been assigned a family. The species Sulfolobus turreted icosahedral virus is so unlike any previously described virus that it will almost certainly be placed in a new family on the next revision of viral families.
Fifteen families are enveloped. These include all three families in the order Herpesvirales and the following families: Ascoviridae, Ampullaviridae, Asfarviridae, Baculoviridae, Fuselloviridae, Globuloviridae, Guttaviridae, Hytrosaviridae, Iridoviridae, Lipothrixviridae, Nimaviridae and Poxviridae.
Bacteriophages (viruses infecting bacteria) belonging to the families Tectiviridae and Corticoviridae have a lipid bilayer membrane inside the icosahedral protein capsid and the membrane surrounds the genome. The crenarchaeal virus Sulfolobus turreted icosahedral virus has a similar structure.
The genomes in this group vary considerably from ~10 kilobases to over 2.5 megabases in length. The largest bacteriophage known is Klebsiella Phage vB_KleM-RaK2 which has a genome of 346 kilobases.
Host range
Species of the order Caudovirales and of the families Corticoviridae and Tectiviridae infect bacteria.
Species of the order Ligamenvirales and the families Ampullaviridae, Bicaudaviridae, Clavaviridae, Fuselloviridae, Globuloviridae and Guttaviridae infect hyperthermophilic archaea species of the Crenarchaeota.
Species of the order Herpesvirales and of the families Adenoviridae, Asfarviridae, Iridoviridae, Papillomaviridae, Polyomaviridae and Poxviridae infect vertebrates.
Species of the families Ascovirus, Baculovirus, Hytrosaviridae, Iridoviridae and Polydnaviruses and of the genus Nudivirus infect insects.
Species of the family Mimiviridae and the species Marseillevirus, Megavirus, Mavirus virophage and Sputnik virophage infect protozoa.
Species of the family Nimaviridae infect crustaceans.
Species of the family Phycodnaviridae and the species Organic Lake virophage infect algae. These are the only known dsDNA viruses that infect plants.
Species of the family Plasmaviridae infect species of the class Mollicutes.
Species of the family Pandoraviridae infect amoebae.
Species of the genus Dinodnavirus infect dinoflagellates. These are the only known viruses that infect dinoflagellates.
Species of the genus Rhizidiovirus infect stramenopiles. These are the only known dsDNA viruses that infect stramenopiles.
Species of the genus Salterprovirus and Sphaerolipoviridae infect species of the Euryarchaeota.
Taxonomy
- Order Caudovirales
- Family Myoviridae â€" includes Enterobacteria phage T4
- Family Podoviridae â€" includes Enterobacteria phage T7
- Family Siphoviridae â€" includes Enterobacteria phage λ
- Order Herpesvirales
- Family Alloherpesviridae
- Family Herpesviridae â€" includes human herpesviruses, Varicella Zoster virus
- Family Malacoherpesviridae
- Order Ligamenvirales
- Family Lipothrixviridae
- Family Rudiviridae
- Unassigned families
- Family Adenoviridae â€" includes viruses which cause human adenovirus infection
- Family Ampullaviridae
- Family Ascoviridae
- Family Asfarviridae â€" includes African swine fever virus
- Family Baculoviridae
- Family Bicaudaviridae
- Family Clavaviridae
- Family Corticoviridae
- Family Fuselloviridae
- Family Globuloviridae
- Family Guttaviridae
- Family Hytrosaviridae
- Family Iridoviridae
- Family Marseilleviridae
- Family Mimiviridae
- Family Nimaviridae
- Family Pandoraviridae
- Family Papillomaviridae
- Family Phycodnaviridae
- Family Plasmaviridae
- Family Polydnaviruses
- Family Polyomaviridae â€" includes Simian virus 40, JC virus, BK virus
- Family Poxviridae â€" includes Cowpox virus, smallpox
- Family Sphaerolipoviridae
- Family Tectiviridae
- Unassigned genera
- Dinodnavirus
- Nudivirus
- Salterprovirus
- Rhizidiovirus
- Unassigned species
- Abalone shriveling syndrome-associated virus
- Bandicoot papillomatosis carcinomatosis virus
- KIs-V
- Haloarcula hispanica pleomorphic virus 1
- Mavirus virophage
- Megavirus
- Organic Lake virophage
- Pithovirus
- Sputnik virophage
- Sputnik virophage 2
- Sulfolobus turreted icosahedral virus
Pleolipoviruses
A group known as the pleolipoviruses, although having a similar genome organisation, differ in having either single or double stranded DNA genomes. Within the double stranded forms have runs of single stranded DNA. This group does not fit into the current classification system and a new taxon is required.
These viruses are nonlytic and form virions characterized by a lipid vesicle enclosing the genome. They do not have nucleoproteins. The lipids in the virla membrane are unselectively acquired from host cell membranes. The virions contain two to three major structural proteins, which either are embedded in the membrane or form spikes distributed randomly on the external membrane surface.
This group includes the following viruses:
- Haloarcula hispanica pleomorphic virus 1
- Halogeometricum pleomorphic virus 1
- Halorubrum pleomorphic virus 1
- Halorubrum pleomorphic virus 2
- Halorubrum pleomorphic virus 3
- Halorubrum pleomorphic virus 6
Group II: ssDNA viruses

Although bacteriophages were first described in 1927, it was only in 1959 that Sinshemer working with phage Phi X 174 showed that they could possess single-stranded DNA genomes. Despite this discovery until relatively recently it was believed that the majority of DNA viruses belonged to the double-stranded clade. Recent work suggests that this may not be the case with single-stranded viruses forming the majority of viruses found in sea water, fresh water, sediment, terrestrial, extreme, metazoan-associated and marine microbial mats. Many of these "environmental" viruses belong to the family Microviridae. However, the vast majority has yet to be classified and assigned to genera and higher taxa. Because most of these viruses do not appear to be related or are only distantly related to known viruses additional taxa will be created for these.
Taxonomy
Families in this group have been assigned on the basis of the nature of the genome (circular or linear) and the host range. Ten families are currently recognised.
- Family Anelloviridae
- Family Bacillariodnaviridae
- Family Bidnaviridae
- Family Circoviridae
- Family Geminiviridae
- Family Inoviridae
- Family Microviridae
- Family Nanoviridae
- Family Parvoviridae
- Family Spiraviridae
Classification
A division of the circular single stranded viruses into four types has been proposed. This division seems likely reflects their phylogenetic relationships.
Type I genomes are characterized by a small circular DNA genome (approximately 2-kb), with the Rep protein and the major open reading frame (ORF) in opposite orientations. This type is characteristic of the circoviruses, geminiviruses and nanoviruses.
Type II genomes have the unique feature of two separate Rep ORFs.
Type III genomes contain two major ORFs in the same orientation. This arrangement is typical of the anelloviruses.
Type IV genomes have the largest genomes of nearly 4-kb, with up to eight ORFs. This type of genome is found in the Inoviridae and the Microviridae.
Given the variety of single stranded viruses that have been described this scheme â€" if it is accepted by the ICTV â€" will need to be extended.
Host range
The families Bidnaviridae and Parvoviridae have linear genomes while the other families have circular genomes. The Bidnaviridae have a two part genome and infect invertebrates. The Inoviridae and Microviridae infect bacteria; the Anelloviridae and Circoviridae infect animals (mammals and birds respectively); and the Geminiviridae and Nanoviridae infect plants. In both the Geminiviridae and Nanoviridae the genome is composed of more than a single chromosome. The Bacillariodnaviridae infect diatoms and have a unique genome: the major chromosome is circular (~6 kilobases in length): the minor chromosome is linear (~1 kilobase in length) and complementary to part of the major chromosome. Members of the Spiraviridae infect archaea.
Molecular biology
All viruses in this group require formation of a replicative form â€" a double stranded DNA intermediate â€" for genome replication. This is normally created from the viral DNA with the assistance of the host's own DNA polymerase.
Recently classified viruses
In the 9th edition of the viral taxonomy of the ICTV (published 2011) the Bombyx mori densovirus type 2 was placed in a new family â€" the Bidnaviridae on the basis of its genome structure and replication mechanism. This is currently the only member of this family but it seems likely that other species will be allocated to this family in the near future.
A new genus â€" Bufavirus â€" was proposed on the basis of the isolation of two new viruses from human stool. These viruses have since been renamed Primate protoparvovirus and been placed in the genus Protoparvovirus.
Unassigned species
A number of additional single stranded DNA viruses have been described but are as yet unclassified.
- Animal viruses - vertebrates
Among these are the parvovirus like viruses. These have linear single stranded DNA genomes but unlike the parvoviruses the genome is bipartate. This group includes the Bombyx mori densovirus type 2, Hepatopancreatic parvo-like virus and Lymphoidal parvo-like virus. A new family Bidensoviridae has been proposed for this group but this proposal has not been ratified by the ICTV to date. Their closest relations appear to be the Brevidensoviruses (family Parvoviridae).
Another new genus â€" as yet unnamed â€" has been proposed. This genus includes the species bovine stool associated circular virus and chimpanzee stool associated circular virus. The closest relations to this genus appear to be the Nanoviridae but further work will be needed to confirm this. Another isolate that appears to be related to these viruses has been isolated from pig faeces in New Zealand. This isolate also appears to be related to the pig stool-associated single-stranded DNA virus. This virus has two large open reading frames one encoding the capsid gene and the other the Rep gene. These are bidirectionally transcribed and separated by intergenic regions. The name Gemycircularvirus has been proposed for this group of viruses. Another virus of this group has been reported again from pigs. Some of this group of viruses may infect fungi. An virus from this group has been isoalted from turkey faeces. Another ten viruses from this group have been isolated from pig faeces.
Fur seal feces-associated circular DNA virus was isolate from the faeces of a fur seal (Arctocephalus forsteri) in New Zealand. The genome has 2 main open reading frames and is 2925 nucleotides in length. Another virus porcine stool associated virus 4 has been isolated that appears to be related to the fur seal virus.
Two viruses have been isolated from human faeces â€" circo-like virus-Brazil hs1 and hs2 â€" with genome lengths of 2526 and 2533 nucleotides respectively. These viruses have four open reading frames. These viruses appear to be related to three viruses previously isolated from waste water, a bat and from a rodent.
Another virus - Porcine stool-associated circular virus 5 - has been reported. This appears to belong to a novel group.
Two viruses have been described from the nesting material yellow crowned parakeet (Cyanoramphus auriceps) â€" CynNCXV (2308 nt) and CynNCKV (2087 nt) Both viruses have two bidirectional open reading frames. Within these are the rolling-circle replication motifs I, II, III and the helicase motifs Walker A and Walker B. There is also a conserved nonanucleotide motif required for rolling-circle replication. CynNCKV has some similarity to the picobiliphyte nano-like virus (Picobiliphyte M5584-5) and CynNCXV has some similarity to the rodent stool associated virus (RodSCV M-45).
Psittacine beak and feather disease virus is a single stranded circular molecule of 1993 nucleotide bases encoding seven open reading frames â€" three in the virion strand and four in the complementary strand. The open reading frames have some homology to porcine circovirus, subterranean clover stunt virus and faba bean necrotic yellows virus.
A virus with a circular genome â€" sea turtle tornovirus 1 â€" has been isolated from a sea turtle with fibropapillomatosis. It is sufficiently unrelated to any other known virus that it may belong to a new family. The closest relations seem to be the Gyrovirinae. The proposed genus name for this virus is Tornovirus.
- Animal viruses - invertebrates
A virus â€" Acheta domesticus volvovirus has been isolated from the house cricket (Acheta domesticus). The genome is circular, has four open reading frames and is 2,517 nucleotides in length. It appears to be unrelated to previously described species. The genus name Volvovirus has been proposed for these species. The genomes in this genus are ~2.5 nucleotides in length and encode 4 open reading frames.
Two new viruses have been isolated from the copepods Acartia tonsa and Labidocera aestiva â€" Acartia tonsa copepod circo-like virus and Labidocera aestiva copepod circo-like virus respectively.
A virus has been isolated from the mud flat snail (Amphibola crenata). This virus has a single stranded circular genome of 2351 nucleotides that encoded 2 open reading frames that are oriented in opposite directions. The smaller open reading frame (874 nucleotides) encodes a protein with similarities to the Rep (replication) proteins of circoviruses and plasmids. The larger open reading frame (955 nucleotides) has no homology to any currently known protein.
An unusual â€" and as yet unnamed â€" virus has been isolated from the flatwom Girardia tigrina. Because of its genome organisation, this virus appears to belong to an entirely new family. It is the first virus to be isolated from a flatworm.
From the hepatopancreas of the shrimp (Farfantepenaeus duorarum) a circular single stranded DNA virus has been isolated. This virus does not appear to cause disease in the shrimp.
A circo-like virus has been isolated from the shrimp (Penaeus monodon). The 1,777-nucleotide genome is circular and single stranded. It has some similarity to the circoviruses and cycloviruses.
Ten new circular viruses have been isolated from dragonfly larvae. The genomes range from 1628 to 2668 nucleotides in length.
- Fungal
Most known fungal viruses have either double stranded DNA or RNA genomes. A single stranded DNA fungal virus â€" Sclerotinia sclerotiorum hypovirulence associated DNA virus 1 â€" has been described. This virus appears to be related to the Geminiviridae but is distinct from them. A genus â€" Breviviridae â€" has been proposed for Sclerotinia sclerotiorum hypovirulence associated DNA virus 1 and a European badger fecal virus.
- Plants
A virus â€" Cassava associated circular DNA virus â€" that has some similarity to Sclerotinia sclerotiorum hypovirulence associated DNA virus 1 has been isolated.
A circular single stranded DNA virus has been isolated from a grapevine. This species may be related to the family Geminiviridae but differs from this family in a number of important respects including genome size.
Grapevine red blotch associated virus and Grapevine cabernet franc associated virus are two single stranded DNA viruses associated with infections of grape vines.
A virus â€" Euphorbia caput medusae latent virus â€" is so divergent from the other members of the geminiviruses that a new genus has been proposed for it. The name of this new genus is proposed to be Capulavirus.
Several viruses â€" baminivirus, nepavirus and niminivirus â€" related to geminvirus have also been reported.
- Archaea
Although ~50 archaeal viruses are known, all but two have double stranded genomes. The first archaeal ssDNA virus to be isolated is the Halorubrum pleomorphic virus 1, which has a pleomorphic enveloped virion and a circular genome.
The second single stranded DNA virus infecting Archaea is Aeropyrum coil-shaped virus (ACV). The genome is circular and with 24,893 nucleotides is currently the largest known ssDNA genome. The viron is nonenveloped, hollow, cylindrical and formed from a coiling fiber. The morphology and the genome appear to be unique. The new family Spiraviridae (from Latin spira, "a coil") has been created by the ICTV to accommodate ACV.
- Marine and other
Several hundred single stranded DNA viral genomes have been isolated from seawater. Their hosts have yet to be identified but are likely to be eukaryotic phytoplankton and zooplankton. They fall into at least 11 distinct groups that are unrelated to previously described viral families.
A virus â€" Boiling Springs Lake virus â€" appears to have evolved by a recombination event between a DNA virus (circovirus) and an RNA virus (tombusvirus). The genome is circular and encodes two proteins â€" a Rep protein and a capsid protein.
Further reports of viruses that appear to have evolved from recombination events between ssRNA and ssDNA viruses have been made.
A new virus has been isolated from the diatom Chaetoceros setoensis. It has a single stranded DNA genome and does not appear to be a member of any previously described group.
Satellite viruses
Satellite viruses are small viruses with either RNA or DNA as their genomic material that require another virus to replicate. There are two types of DNA satellite viruses â€" the alphasatellites and the betasatellites â€" both of which are dependent on begomaviruses. At present satellite viruses are not classified into genera or higher taxa.
Alphasatellites are small circular single strand DNA viruses that require a begomovirus for transmission. Betasatellites are small linear single stranded DNA viruses that require a begomovirus to replicate.
Phylogenetic relationships

Introduction
Phylogenetic relationships between these families are difficult to determine. The genomes differ significantly in size and organisation. Most studies that have attempted to determine these relationships are based either on some of the more conserved proteins â€" DNA polymerase and others â€" or on common structural features. In general most of the proposed relationships are tentative and have not yet been used by the ICTV in their classification.
ds DNA viruses
- Herpesviruses and caudoviruses
While determining the phylogenetic relations between the various known clades of viruses is difficult, on a number of grounds the herpesviruses and caudoviruses appear to be related.
While the three families in the order Herpesvirales are clearly related on morphological grounds, it has proven difficult to determine the dates of divergence between them because of the lack of gene conservation. On morphological grounds they appear to be related to the bacteriophages â€" specifically the Caudoviruses.
The branching order among the herpesviruses suggests that Alloherpesviridae is the basal clade and that Herpesviridae and Malacoherpesviridae are sister clades. Given the phylogenetic distances between vertebrates and molluscs this suggests that herpesviruses were initially fish viruses and that they have evolved with their hosts to infect other vertebrates.
The vertebrate herpesviruses initially evolved ~400 million years ago and underwent subsequent evolution on the supercontinent Pangaea. The alphaherpesvirinae separated from the branch leading to the betaherpesvirinae and gammaherpesvirinae about 180 million years ago to 220 million years ago. The avian herpes viruses diverged from the branch leading to the mammalian species. The mammalian species divided into two branches â€" the Simplexvirus and Varicellovirus genera. This latter divergence appears to have occur around the time of the mammalian radiation.
Several dsDNA bacteriophages and the herpesviruses encode a powerful ATP driven DNA translocating machine that encapsidates a viral genome into a preformed capsid shell or prohead. The critical components of the packaging machine are the packaging enzyme (terminase) which acts as the motor and the portal protein that forms the unique DNA entrance vertex of prohead. The terminase complex consists of a recognition subunit (small terminase) and an endonuclease/translocase subunit (large terminase) and cuts viral genome concatemers. It forms a motor complex containing five large terminase subunits. The terminase-viral DNA complex docks on the portal vertex. The pentameric motor processively translocates DNA until the head shell is full with one viral genome. The motor cuts the DNA again and dissociates from the full head, allowing head-finishing proteins to assemble on the portal, sealing the portal, and constructing a platform for tail attachment. Only a single gene encoding the putative ATPase subunit of the terminase (UL15) is conserved among all herpesviruses. To a lesser extent this gene is also found also in T4-like bacteriophages suggesting a common ancestor for these two groups of viruses.
A common origin for the herpesviruses and the caudoviruses has been suggested on the basis of parallels in their capsid assembly pathways and similarities between their portal complexes, through which DNA enters the capsid. These two groups of viruses share a distinctive 12-fold arrangement of subunits in the portal complex.
- Large DNA viruses
The family Ascoviridae appear to have evolved from the Iridoviridae. The family Polydnaviridae may have evolved from the Ascoviridae. Molecular evidence suggests that the Phycodnaviridae may have evolved from the family Iridoviridae. These four families (Ascoviridae, Iridoviridae, Phycodnaviridae and Polydnaviridae) may form a clade but more work is needed to confirm this.
Based on the genome organisation and DNA replication mechanism it seems that phylogenetic relationships may exist between the rudiviruses (Rudiviridae) and the large eukaryal DNA viruses: the African swine fever virus (Asfarviridae), Chlorella viruses (Phycodnaviridae) and poxviruses (Poxviridae).
Based on the analysis of the DNA polymerase the genus Dinodnavirus may be a member of the family Asfarviridae. Further work on this virus will required before a final assignment can be made.
The nucleocytoplasmic large DNA virus group (Asfarviridae, Iridoviridae, Marseilleviridae, Mimiviridae, Phycodnaviridae and Poxviridae) along with three other families â€" Adenoviridae, Cortiviridae and Tectiviridae â€" and the phage Sulfolobus turreted icosahedral virus and the satellite virus Sputnik all possess double β-barrel major capsid proteins suggesting a common origin.
Some of the relations among the large viruses have been established. Mimiviruses are distantly related to Phycodnaviridae. Pandoraviruses share a common ancestor with Coccolithoviruses within the Phycodnaviridae family. Pithoviruses are related to Iridoviridae and Marseilleviridae.
- Other viruses
Based on the analysis of the coat protein, Sulfolobus turreted icosahedral virus may share a common ancestry with the Tectiviridae.
The families Adenoviridae and Tectiviridae appear to be related structurally.
Baculoviruses evolved from the nudiviruses 310 million years ago.
The Hytrosaviridae are related to the baculoviruses and to a lesser extent the nudiviruses suggesting they may have evolved from the baculoviruses.
The Nimaviridae may be related to nudiviruses and baculoviruses.
The Nudiviruses seem to be related to the polydnaviruses.
A protein common to the families Bicaudaviridae, Lipotrixviridae and Rudiviridae and the unclassified virus Sulfolobus turreted icosahedral virus is known suggesting a common origin.
Examination of the pol genes that encode the DNA dependent DNA polymerase in various groups of viruses suggests a number of possible evolutionary relationships. All know viral DNA polymerases belong to the DNA pol families A and B. All possess a 3'-5'-exonuclease domain with three sequence motifs Exo I, Exo II and Exo III. The families A and B are distinguishable with family A Pol sharing 9 distinct consensus sequences and only two of them are convincingly homologous to sequence motif B of family B. The putative sequence motifs A, B, and C of the polymerase domain are located near the C-terminus in family A Pol and more central in family B Pol.
Phylogenetic analysis of these genes places the adenoviruses (Adenoviridae), bacteriophages (Caudovirales) and the plant and fungal linear plasmids into a single clade. A second clade includes the alpha- and delta-like viral Pol from insect ascovirus (Ascoviridae), mammalian herpesviruses (Herpesviridae), fish lymphocystis disease virus (Iridoviridae) and chlorella virus (Phycoviridae). The pol genes of the African swine fever virus (Asfarviridae), baculoviruses (Baculoviridae), fish herpesvirus (Herpesviridae), T-even bacteriophages (Myoviridae) and poxviruses (Poxviridae) were not clearly resolved. A second study showed that poxvirus, baculovirus and the animal herpesviruses form separate and distinct clades. Their relationship to the Asfarviridae and the Myoviridae was not examined and remains unclear.
The polymerases from the archaea are similar to family B DNA Pols. The T4-like viruses infect both bacteria and archaea and their pol gene resembles that of eukaryotes. The DNA polymerase of mitochondria resembles that of the T odd phages (Myoviridae).
The virophage â€" Mavirus â€" may have evolved from a recombination between a transposon of the Polinton (Maverick) family and an unknown virus.
ss DNA viruses
The evolutionary history of this group is currently poorly understood. An ancient origin for the single stranded circular DNA viruses has been proposed.
Capsid proteins of most icosahedral ssRNA and ssDNA viruses display the same structural fold, the eight-stranded beta-barrel, also known as the jelly-roll fold. On the other hand, the replication proteins of icosahedral ssDNA viruses belong to the superfamily of rolling-circle replication initiation proteins that are commonly found in prokaryotic plasmids. Based on these observations, it has been proposed that small DNA viruses have originated via recombination between RNA viruses and plasmids.
Circoviruses may have evolved from a nanovirus.
Given the similarities between the rep proteins of the alphasatellites and the nanoviruses, it is likely that the alphasatellites evolved from the nanoviruses. Further work in this area is needed to clarify this.
The geminiviruses may have evolved from phytoplasmal plasmids.
Based on the three-dimensional structure of the Rep proteins the geminiviruses and parvoviruses may be related.
The ancestor of the geminiviruses probably infected dicots.
The parvoviruses have frequently invaded the germ lines of diverse animal species including mammals, fishes, birds, tunicates, arthropods and flatworms. In particular they have been associated with the human genome for ~98 million years.
Bacteriophage evolution
Since 1959 ~6300 prokaryote viruses have been described morphologically, including ~6200 bacterial and ~100 archaeal viruses. Archaeal viruses belong to 15 families and infect members of 16 archaeal genera. These are nearly exclusively hyperthermophiles or extreme halophiles. Tailed archaeal viruses are found only in the Euryarchaeota, whereas most filamentous and pleomorphic archaeal viruses occur in the Crenarchaeota. Bacterial viruses belong to 10 families and infect members of 179 bacterial genera: most these are members of the Firmicutes and γ-proteobacteria.
The vast majority (96.3%) are tailed with and only 230 (3.7%) are polyhedral, filamentous or pleomorphic. The family Siphoviridae is the largest family (>3600 descriptions: 57.3%). The tailed phages appear to be monophyletic and are the oldest known virus group. They arose repeatedly in different hosts and there are at least 11 separate lines of descent.
All of the known temperate phages employ one of only three different systems for their lysogenic cycle: lambda-like integration/excision, Mu-like transposition or the plasmid-like partitioning of phage N15.
A putative course of evolution of these phages has been proposed by Ackermann.
Tailed phages originated in the early Precambrian, long before eukaryotes and their viruses. The ancestral tailed phage had an icosahedral head of about 60 nanometers in diameter and a long non contractile tail with sixfold symmetry. The capsid contained a single molecule of double stranded DNA of about 50 kilobases. The tail was probably provided with a fixation apparatus. The head and tail were held together by a connector. The viral particle contained no lipids, was heavier than its descendant viruses and had a high DNA content proportional to its capsid size (~50%). Most of the genome coded for structural proteins. Morphopoietic genes clustered at one end of the genome, with head genes preceding tail genes. Lytic enzymes were probably coded for. Part of the phage genome was nonessential and possibly bacterial.
The virus infected its host from the outside and injected its DNA. Replication involved transcription in several waves and formation of DNA concatemers.
New phages were released by burst of the infected cell after lysis of host membranes by a peptidoglycan hydrolase. Capsids were assembled from a starting point, the connector and around a scaffold. They underwent an elaborate maturation process involving protein cleavage and capsid expansion. Heads and tails were assembled separately and joined later. The DNA was cut to size and entered preformed capsids by a headful mechanism.
Subsequently the phages evolved contractile or short tails and elongated heads. Some viruses become temperate by acquiring an integrase-excisionase complex, plasmid parts or transposons.
NCLDVs

The asfarviruses, iridoviruses, mimiviruses, phycodnaviruses and poxviruses have been shown to belong to a single group, â€" the large nuclear and cytoplasmic DNA viruses. These are also abbreviated "NCLDV". This clade can be divided into two groups:
- the iridoviruses-phycodnaviruses-mimiviruses group. The phycodnaviruses and mimiviruses are sister clades.
- the poxvirus-asfarviruses group.
It is probable that these viruses evolved before the separation of eukaryoyes into the extant crown groups. The ancestral genome was complex with at least 41 genes including (1) the replication machinery (2) up to four RNA polymerase subunits (3) at least three transcription factors (4) capping and polyadenylation enzymes (5) the DNA packaging apparatus (6) and structural components of an icosahedral capsid and the viral membrane.
The evolution of this group of viruses appears to be complex with genes having been gained from multiple sources.
Another group of large viruses â€" the Pandoraviridae â€" has been described. Two species â€" Pandoravirus salinus and Pandoravirus dulcis â€" have been recognized. These were isolated from Chile and Australia respectively. These viruses are about one micrometer in diameter making them one of the largest viruses discovered so far. Their gene complement is larger than any other known virus to date. At present they appear to be unrelated to any other species of virus.
An even larger genus, Pithovirus, has since been discovered, measuring about 1.5 µm in length.
References
Additional reading

- Fauquent, C. M.; Mayo, M. A.; Maniloff, J.; Desselberger, U.; Ball, L. A.; eds. (2005-05-27). "8th Reports of the International Committee on Taxonomy of Viruses". Academic Press. U.S. National Center for Biotechnology Information, National Library for Medicine, National Institutes of Health.Â





I'm 61 years old, I contracted hpv in 2011' I has be taking lot treatment for it and embarrassedsome months ago the wart stated coming out seriously, I used lot recommendation because there was lot warts around my anus and was so . but today I'm totally happy I got the virus eliminated by using natural treatment from Dr Onokun herbal center after his treatment I got cured. all the warts went away' seriously believed Dr Onokun he have the cure for human papillomavirus because he has eliminated hpv been in my body since 2011, Dr Onokun make it possible for me. Here is Dr Onokun email to reach him: Dronokunherbalcure@gmail.com he is welled capable of curing terrible diseases.
ReplyDeleteI has suffered for Human papillomavirus HPV) for 2years, I was given some tablets at the hospital but I refused to take it, They said I have to be on it for life so I don't want take a drugs everyday for life. No point in taking medicine everyday when u won't get cure from it and I was advice to seek for natural herbal cure, after some time I found dr onokun is the most trustful herbalist that have herbs to cure wicked symptom's,I emailed dr onokun, for 2weeks been his patient he cured my (HPV) with his herbal. I only used his natural herbs for two weeks it was 100% cure. I'm not (HPV) patient anymore. I'm happy about it i finally got cured out of this mess been in my body for 2years. I also recommend you if you're living with (HPV) or herpes symptoms i also want you to be free contact dr onokun with the email attach to my post. Dronokunherbalcure@gmail.com or website: www.dronokunherbalcure.wordpress.com
ReplyDeleteI found out, that taking herbal treatment is the best to get rid of hpv as soon as i feel the symptoms of hpv is appearing, i took the healing process by contacting Dr onokun for natural treatment it works wonders, amazingly ever since I had the herbal treatment i have not feel these horrible disease anymore and my doc told me the virus is gone, i am glad i finally got cured out from this horrible disease. every hpv or herpes patients should also get in touch with this herbalist Dr to get ride of these disease forever his email address; Dronokunherbalcure@gmail.com
ReplyDelete